Memory: human capacity to encode, store, and retrieve past experience
Memory is the cognitive capacity to register, retain, and recall information and experience; it has multiple types, neural mechanisms, and practical uses in daily life and science.
Memory is the mental capacity to register, retain and later retrieve information about past experiences. It depends on the functioning of the brain and on processes that allow information to be transformed into a stable form for later use. Memory makes it possible to recognize people and places, learn skills, plan actions, and maintain a continuous sense of identity over time.
Image gallery
9 Images






Types and characteristics
Researchers usually distinguish several interacting forms of memory rather than a single thing. Major categories include:
- Sensory memory: brief traces of incoming sensory input.
- Short-term and working memory: temporary holding and manipulation of information for current tasks; often called working memory.
- Long-term memory: durable storage that can be subdivided into episodic (events), semantic (facts and concepts), and procedural (skills and habits) types.
Key processes are encoding (how experiences are converted into memory), storage (maintenance over time), and retrieval (accessing stored information). Effective encoding often requires attention, meaningful connections, and repetition; techniques such as mnemonics or spaced practice improve retention.
Neural mechanisms and study
Memory arises from patterns of neural activity and lasting changes in synaptic connections. Structures such as the hippocampus play an important role in forming new declarative memories, while distributed cortical networks support long-term storage. At the cellular level, phenomena like synaptic plasticity and long-term potentiation are widely studied as mechanisms that strengthen connections after learning. The scientific study of memory is a core topic in cognitive psychology and intersects neuroscience, biology, and computer science.
Typical everyday memory can be described simply as the brain's ability to store information for later use. This capacity varies across individuals and across the lifespan: development in childhood improves certain memory skills, while aging and neurological conditions can impair them. Disorders such as amnesia and Alzheimer-type dementia illustrate how memory loss affects functioning and personality.
History and cultural views shape how memory is valued. Ancient Greeks personified memory as Mnemosyne (Mnemosyne), reflecting its central role in art and knowledge. Today memory research informs education, clinical treatment, legal practices (eyewitness testimony), and technology (artificial memory models). Simple strategies—adequate sleep, focused practice, retrieval practice, and spaced repetition—remain among the most reliable ways to support healthy memory.
Notable distinctions include the difference between remembering facts versus performing skills, conscious versus implicit remembering, and transient forgetting versus permanent loss. Understanding these distinctions helps in designing learning methods, diagnosing memory disorders, and creating supportive environments for people across the lifespan.
Classification into different types of memory
Memory can be divided into different subsystems according to the duration of information storage. Usually three systems are distinguished:
- Sensory memory (also sensory register): Holds information for milliseconds to seconds.
- Working memory (also short-term memory): stores information for about 20-45 seconds
- Long-term memory: Stores information for years
A different model is represented by the levels-of-processing approach.
Sensory memory (ultra-short-term memory)
New information reaches the brain via the sensory organs and is temporarily stored in sensory memory (also called sensory register, formerly also called immediacy memory, ultra-short-term memory or ultra-short-term memory). Sensory memory is specific to each sensory modality; it is also called iconic memory for visual perception and echoic memory for auditory perception. The ability to repeat something previously said in a conversation, despite not having just listened, is an example of auditory sensory memory.
Far more information is recorded in sensory memory than in working memory. However, this information decays after only a few tenths of a second. One way of investigating the decay of information in this memory system is the so-called partial-report method, which was developed by George Sperling (1960). In this method, test subjects are presented with several series of letters (sets) in different lines, of which only individual lines are to be reproduced during subsequent recall. This is intended to prevent the others from being forgotten during the time when individual parts of the set are recalled. If in an experiment the time between the presentation of the set and the indication of which line is to be played back is varied and the memory performance is compared according to the intermediate time, an estimate of the duration of storage is obtained. Using this method, it has been shown that visual sensory memory can store information for about 15 milliseconds, whereas auditory sensory memory can store information for about 2 seconds.
In this type of memory, centrally controlled processes such as consciousness or attention usually do not play a significant role. However, these can have a major influence in the transfer of information to working memory.
Short-term memory and working memory
The basis of conscious information processing is short-term memory (in some models also working memory). Short-term memory is a store that holds a narrowly limited amount of information in an immediately available state.
According to a hypothesis now considered historically outdated, it had an approximate capacity of about 7 ± 2 units of information, as long as it was numerically listable things. These were also called chunks (see there for more recent findings).
Short-term memory
One aspect that has received particular attention in the study of short-term memory is "rapid forgetting". This was first investigated by Peterson & Peterson (1959). By showing their subjects single words, word triads, and consonant triads followed by a distracting task (counting backwards), they found a significant drop in memory performance as a function of the length of the distracting task. In addition, it made a difference whether the words were presented individually or in groups. Single words showed a significantly lower forgetting rate than a group of three consonants or three words. The latter two did not differ from each other. Murdock (1961) confirmed the results of Peterson & Peterson and could additionally show that the presentation of several items of the same semantic category caused a forward inhibition. The subjects found it more difficult to distinguish between things the more they had seen (list length effect). This was reflected in a marked drop in memory performance.
Delos Wickens (1970) was able to show that forward inhibition could be reversed when subjects were presented with words of different semantic categories. After a category change, memory performance increased again significantly. Gunter et al. (1981) conducted three experiments in which they demonstrated forward inhibition and its reversal. They had their subjects recite individual television news stories of different subject areas, ranging from, for example, domestic and foreign policy topics. One group was presented with four similar topics, the other with three similar topics and one news item from a different topic area. The first group showed forward inhibition in terms of decreasing memory performance, and the second group showed inhibition reversal in terms of changing topics. Both effects were also found with a reduced number of items and with the additional task of describing them accurately. In addition, the authors were able to demonstrate a learning effect when certain things had already been shown in a previous test. The subjects were then better able to remember them in a second test. Investigations into the time period of the effect of forward inhibition pointed most likely to the retrieval phase.
Working memory
The original model of short-term memory has been supplemented since 1974 by Baddeley's working memory model, which cites the following three systems:
- The spatial-visual notepad for short-term storage of visual impressions.
- The articulatory or phonological loop is used to store verbal information, which can remain available for a relatively long time through internal repetition.
- The central executive manages the two subsystems and links information from them to long-term memory.
Most recently, an episodic buffer has been added to the model.
Long-term memory
Long-term memory is the brain's permanent storage system. It is not a single entity, but multiple storage services for different types of information. It can be stored in long-term memory from minutes to years (secondary memory) or even for a lifetime (tertiary memory). Nothing is known about limitations of the capacity of long-term memory. However, studies in so-called savants (French) or insular gifted individuals suggest a significantly higher memory capacity than that used normally. Forgetting does not seem to be a capacity problem, but a protection against too much knowledge. Forgetting seems to take place less by loss of information as in the other, short-term forms of memory, but by deleting or falsifying influence of other, before or after formed contents.
Different processes of the long-term memory are to be distinguished:
- Learning/encoding: new storage of information
- Remembering/Recalling: Becoming aware of memory contents
- Consolidation/Retention: Consolidation of information through repeated recall
- Linking new and old information
- Forgetting: Decay of memory content or alteration by competing information.
For the transfer of new memory contents into the long-term memory and the retention of information, practice is often beneficial, for example, through the conscious recall and reconsideration of information in the working memory. Anchorage in memory increases with importance, emotional weight and the number of associations (linking with other content).
A single stored and retrievable piece of information is called an engram (memory trace). The totality of all engrams forms the memory.
Components of long-term memory
Basically, two forms of long-term memory are distinguished, which store different types of information: Declarative (explicit) and procedural implicit memory. The different forms of information are independent of each other and are stored in different areas of the brain, so that, for example, patients with amnesia (memory disorder) of the declarative memory can have undisturbed procedural memory.
Declarative memory
Declarative memory", also known as knowledge memory, stores facts and events that can be consciously recalled. Declarative memory is divided into two areas:
- The "semantic memory" contains the world knowledge, general facts independent of the person ("Paris is the capital of France", "One has a mother and a father").
- Episodic memory" contains episodes, events and facts from one's own life (memory of experiences during a visit to Paris, the face and name of one's own father).
Procedural memory
Procedural memory, also known as behavioral memory, stores automated sequences of actions or skills. Examples are walking, cycling, dancing, driving, playing the piano. These are complex movements whose sequence has been learned and practiced and which are then recalled and executed without thinking.
Memory capacity
The capacity of human memory is difficult to determine and depends on the type of information we store. For example, it has been estimated that on average each person can recognize and thus remember about 5000 faces of other people.
Anatomy and physiology of memory
Unlike other areas such as language, motor skills, vision or hearing, there is no definable comprehensive "memory area" in the brain. Rather, memory is predominantly based on additional services provided by parts of the brain that are specialized in other ways. Nevertheless, different anatomical structures can be distinguished that are necessary for memory. First of all, it must be clarified what represents the correlate (correspondence) of learning and memory at the lowest level, at the individual neuron.
Neuronal learning processes
The memory content is laid down in the connections of the nerve cells, the synapses, more precisely in the synaptic efficiency of neuronal networks. After the hypothesis was held until the 1970s that chemical molecules could take over this role - scotophobin has become particularly famous - this hypothesis turned out to be no longer tenable.
There are an estimated 100 to 500 trillion synapses between the approximately 100 billion nerve cells. The key factor here is synaptic plasticity: many synapses are anatomically adaptable. This allows them to alter the efficiency of transmission between neurons. In addition, transmission properties are adapted by the formation and degradation of new synapses.
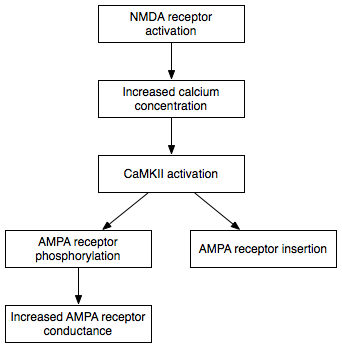
In 1949, Donald O. Hebb was the first to propose that synapses - depending on the extent of their activation by neuronal activity - "permanently" change the strength of their signaling ability by anatomical remodeling. The hypothesis he put forward in what he called Hebb's learning rule has been experimentally confirmed. Thus, a synapse that becomes stronger as a result of simultaneous activity in the pre- and postsynaptic neuron is called a "Hebb synapse". Such a permanent change of a synapse is called "homosynaptic" long-term potentiation (long-term strengthening) in neurophysiology.
There are a variety of other forms of synaptic plasticity. They differ mainly in their direction (potentiation or depression, i.e. strengthening or weakening), in their duration (short-term or long-term change), in their synaptic specificity (homo- or heterosynaptic), and in the molecular mechanisms of their development and maintenance.
Various signalling cascades have been described that start with the excitation of a nerve cell by a specific synapse and a subsequent action potential and lead to short- or long-term changes in synaptic efficiency. Such mechanisms include, in the short term, the phosphorylation of receptor molecules, the release of retrograde messengers for the presynaptic axon (nerve fiber), and, for the long term, in particular the activation of transcription factors that regulate protein biosynthesis and lead to increased synthesis of receptor molecules, enzymes for transmitter uptake and degradation, and structural proteins.
Anatomical coarse structures
Today, specific brain regions are assigned to the different types of memory. The assignments could be made by comparing memory disorders in localized damage to the brain (for example, due to stroke).
Working memory is assigned to the prefrontal cortex. Long-term memory, on the other hand, is based on the interaction of the cortex and numerous subcortical areas. A distinction is made between the different qualities of information.
Declarative memory
The entire neocortex is involved in declarative memory, the right frontal and temporal cortex in particular in episodic memory, and the temporal lobe in particular in semantic memory.
However, subcortical regions, such as the limbic system, especially the medial temporal lobe system, the hippocampus and adjacent areas, are also involved, particularly in the process of storage. These are grouped together in the so-called Papez neuron circuit. Often cited is the case of patient HM, who had both hippocampi removed for therapy of severe epilepsy. Although the epilepsy was cured, the patient showed severe anterograde amnesia after the operation: he could not remember anything new. In contrast, access to memory content acquired before the operation was not impaired.
Procedural memory
In addition to cortex areas such as the motor and prefrontal areas, the cerebellum and the basal ganglia are particularly involved in skill learning in humans. The amygdala plays an important role in the storage of emotionally significant memory content, as well as fear reactions.
For forms of learning in the manner of classical conditioning, which are also present in more primitive animals, evolutionarily older brain areas are accordingly also involved. Often, the place of learning here is where the two stimuli to be linked anatomically converge. In particular, the cerebellum plays a role here.


Related articles
Author
AlegsaOnline.com Memory: human capacity to encode, store, and retrieve past experience Leandro Alegsa
URL: https://en.alegsaonline.com/art/63750
Sources
- books.google.com : AS Level Psychology Through Diagrams