Language: human communication, structure, history and uses
An overview of human language: its nature, structure (syntax, words, rules), modalities (speech, writing, sign), history and change, social roles, and threats such as language endangerment.
Overview. Language is the primary system by which humans communicate complex ideas, feelings and information. The scientific study of this faculty is called linguistics. Human languages combine sounds, gestures or symbols with conventional meanings, allowing abstract thought and coordinated action beyond the capabilities of typical animal signals.
Image gallery
10 Images









Structure and components
At its core, language organizes meaningful units into larger patterns. Phonology and morphology build and shape sounds or signs into words; syntax governs how those words are combined by rules to make statements and questions. Semantics treats meaning and pragmatics studies how context affects interpretation. These interacting levels let speakers create novel sentences and convey subtle meanings.
Modalities and expression
Language is not confined to vocal speech: it can be realized in different channels. Common modalities include speech, conventionalized writing, and visual-gestural systems such as sign language. Each modality has its own constraints and advantages for memory, transmission and social use. People also use language internally for thinking, planning and imagination.
Origins, change and formalizations
How language arose in human evolution remains debated: proposals draw on comparative primate behavior, archaeology and cognitive science. Over historical time languages diverge, borrow and create new vocabulary to describe inventions and social change. Some systems have been abstracted into formal language frameworks used in mathematics and logic, or encoded for computing, showing how natural language inspires precise symbolic systems.
Social roles and varieties
Languages function at the level of a community and often serve as markers of identity for a country or group. They appear in different registers (formal, colloquial), dialects and specialized jargons. Language study is also part of education as a school subject, where learners acquire literacy and cultural knowledge.
Importance and challenges
Language enables science, law, literature and everyday cooperation, but many languages face decline. Organizations such as UNESCO document and support endangered traditions: thousands of tongues are considered endangered as communities shift to dominant languages. Preserving linguistic diversity matters for cultural heritage, ecological knowledge and cognitive science.
- Key features: productivity, displacement, arbitrariness.
- Common modalities: speech, writing, sign language.
- Related disciplines: linguistics, anthropology, cognitive science.
Language as a sign system
Individual languages
→ Main article: Individual language and language family
In a specific sense, the word language refers to a particular individual language such as German, Japanese or Swahili. The spoken languages of mankind are divided into language families according to their genetic relationship. Each individual language is unambiguously classified internationally on the basis of the so-called language codes according to the ISO-639 sub-standards. Of the approximately 6,500 individual languages counted today - according to the National Geographic Society, as many as 6,912 languages were actively used worldwide in 2005 - more than half are threatened by language death, since they are hardly ever spoken and are often no longer passed on to children. It is therefore assumed that a large proportion of the languages still in existence today will disappear within the next 100 years. Currently, the most common 50 languages are spoken by about 80 percent of humanity as a mother tongue (and by about 90 % also as a second language), all other (still) existing languages by the remaining 20 percent of people.
The demarcation of language and dialect was very controversial for a long time, but today, according to the theories of Heinz Kloss, it is almost always possible on the basis of scientific criteria (see also the article Dialect).
From an ethnological and sociolinguistic point of view, the languages used by humans in everyday life are divided in terms of their origin into naturally evolved languages such as English or Spanish and deliberately constructed languages such as Esperanto or Klingon.
Even natural language can be influenced by conscious planning, as in the case of German in Martin Luther's translation of the Bible. This variety was a constructed form that was to be understood everywhere. As a result, it eventually became a lingua franca. Variants of ethnic languages are sometimes unified into a variety by design in the course of language policy measures, as in the case of Ladin in South Tyrol/Northern Italy. However, there are limits to the controllability of natural language, since linguistic knowledge is unconscious knowledge, which is also acquired very early in life and unconsciously. Every natural language shows historical change in this context, which often overrides attempts at standardization.
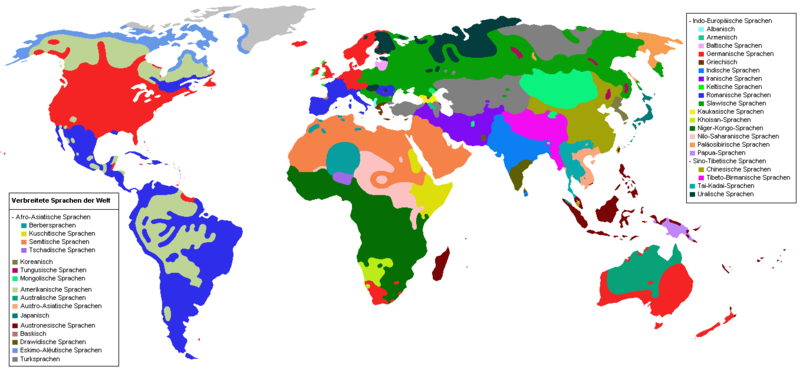
enlarge and show information about the picture
![]()
Language families of the world
Constructed and formal languages
→ Main article: Formal language
Unlike single natural languages, formal languages can be described by logic and set theory (enumerable set of basic expressions, clear rules of composition, well-formed expressions). They are used, for example, in theoretical computer science, especially in computability theory and compiler construction. Programming languages such as ALGOL, APL, Fortran, COBOL, BASIC, C, C++, Ada, Lisp, Prolog, Python, Java or Perl are constructed for specific purposes and are based on theoretical as well as pragmatic considerations.
With his project of the ortho-language program, the mathematician Paul Lorenzen pursued the construction of an unambiguous and methodically structured scientific language, which, however, was "highly controversial even in methodological philosophy".
The descriptive principles of formal logic are also applied to natural language; pioneering work on this was done by the American logician Richard Montague.
The human spoken language as a sign system
Human spoken language can also be understood as a sign system (see semiotics), consisting of a multitude of signs that have a meaning (see semantics), which can be linked by means of grammatical rules (syntactics) to form an infinite number of statements. Ferdinand de Saussure conceived the linguistic sign as an arbitrary, non-compulsory combination of phonetic image (signifiant = the signified) and mental image (signifié = the signified).
Humboldt's remark about the convention theory of language reads like a critique of 20th century semiotics:
"The most detrimental influence on the interesting treatment of every study of language has been exerted by the limited notion that language originated by convention, and that the word is nothing but the sign of a thing, or of a concept, existing independently of it. This view, which is undeniably correct up to a certain point, but which is also quite wrong beyond that point, kills all spirit and banishes all life as soon as it begins to prevail, and it is to it that one owes the commonplaces so often repeated: [...] that every language, if one only knows how to use it properly, is about equally good [...] language is a separate and independent being, an individual, the sum of all words, language, is a world that lies in the middle between the one that appears outside, and the one that acts within us [...]"
That is, the meaning of words is developed in living dialogue; language suspends the reality of the object and the subject within itself, it mediates between the two, and so in language, beyond convention, "every new [view] can always discover something new." Even in the case of empirical objects, the words of different languages are never perfect synonyms; all the more so in the case of designations for thoughts and sensations with even more indeterminate outlines.
Evolution of language ability
→ Main article: Language origin and development
Language as an "instinct to learn
Already Darwin distinguished between the biological ability of man, which allows him to acquire language, and specific languages as such. This theoretical distinction is adopted by modern cognitive biology. Babies have an instinct to babble, but need to learn language. Thus, for ethologist Peter Marler, as for Darwin, language was not an instinct, but "language is an instinct to learn, the expression of which implies that both biological and external conditions are met." It is to this "instinct" to learn language that the biological evolutionary study of language ability is directed. An important language-related gene discovered in this setting is FOXP2, a phylogenetically ancient transcription factor that plays a role in flexible oral-motor vocal control. FOXP2 underwent a crucial mutation in the human genus at least 400,000 years ago, which is inferred from Neanderthal having the same allele. A set of four characteristic genes has been identified for simple aspects of syntax.
Anatomy
Until around 2010, the prevailing view on the evolutionary language ability of humans was that anatomically modern humans (Homo sapiens) differed from apes in their ability to speak. According to this view, language rich in variation only became possible through anatomical changes in the course of the phylogeny of humans. It is unknown how developed the ability to speak was in the common ancestor of Neanderthal man and Homo sapiens, Homo erectus. Likewise, how "advanced" the morphological and functional potential for differentiated linguistic communication was in the transition from Homo erectus to early anatomically modern humans is unknown. The enlargement of the pharyngeal cavity (as a resonating body), the lowering of the larynx, and the bulging of the palate, which had already begun in Homo erectus, were seen as necessitating greater freedom of movement for the tongue. In the interaction of the pharyngeal cavity, the oral and nasal cavities, the soft palate, the lips and the tongue, the fundamental tone produced by the vocal cords can then be modulated into vowels and consonants. Skull findings prove that the arching of the palate and the lowering of the larynx were completed about 100,000 years ago.
In the Kebara Cave near Haifa in Israel, a hyoid bone was found in a skeleton of a Neanderthal man that was about 60,000 years old, suggesting that this man was capable of spoken language. Anthropologists from Durham suggest that Neanderthal ancestors were able to speak more than 300,000 years ago. They compared the size of the "canalis nervi hypoglossi," an opening in the base of the skull, in skulls of modern humans with various fossils. According to these anthropologists, a large hypoglossal nerve is a prerequisite for differentiated speech. Through this opening at the base of the skull runs the nerve by which the brain controls the movement of the tongue. Scientists found that the canalis nervi hypoglossi was similar in size in Neanderthals as it is in modern humans. In pre-humans of the genus Australopithecus, who lived about two million years ago, it is much smaller.
Recent research results show that the lowering of the larynx was not a solely human characteristic, but occurred in the animal kingdom in many cases, for example in red deer or wapiti deer. At the same time, the previously denied dynamics and reconfigurability of the vocal range is now confirmed in empirical studies for animals, for example in many mammals such as dogs, goats, seals, and also in alligators. Because of the phylogenetically different ancestry of the species examples mentioned, it is assumed that the lowering of the larynx was an evolutionarily early feature. The reasons for this may lie, for example in the deer, in sexual selection through lowered vocalization. The ability to learn song is also inherent in birds. These findings imply, first, that the vocal tract was sufficiently flexible for complex language development at any point in primate evolution and, second, that fossil evidence from human ancestors provides little evidence for language ability. The evolutionary prerequisites for language are now seen more in neurological control or mechanisms and less in the anatomy of the vocal tract.
Neural prerequisites
Whereas language used to be treated as a monolithic entity, cognitive biology today breaks down cognitive language prerequisites into separable components and analyzes them comparatively in different animal strains. The following are seen as prerequisites for the evolution of language: social intelligence, imitation, eye contact sensitivity, spatial gaze-following ability, and the Theory of Mind. These mechanisms form core elements of animal social behavior. Our ability to share thoughts socially allows human cultures to accumulate knowledge in ways that would not be possible without language. Preliminary stages of language have been empirically explored in recent years. According to current research, no evolutionary linear higher development of language exists as animal phyla converge with humans. In birds, similar cognitive, presuppositional abilities are seen in tests as in primates.
Protolanguage Models
Language evolution explores models of protolanguages. Protolanguage differs from protolanguage and refers to alternative forms of communication (models) from which the protolanguage, if it existed, could first emerge. Three models are distinguished: the lexical model, the gestural model and the musical model. All models should provide answers to three components of language, signals, syntax and semantics. These components can be considered as key evolutionary innovations that have evolved since humans split from the last common ancestor. The lexical protolanguage contained spoken words. Syntax as an innovation came later, its emergence, especially in terms of multiple semantic hierarchies, is unclear, as are still the cognitive mechanisms to unambiguously interpret word meaning in the context of language when words are ambiguous. Situationally changing alarm calls of the southern green monkey can be considered an example of a primordial state of lexical protolanguage, but the calls are not learned in the sense of language learning. The lexical protolanguage also does not have the property of intention for information transmission. The gestural model assumes that language arose from pointing gestures. Sign language today can be a complete language with syntax and semantics. Great apes are better at pointing gestures than language. The question then becomes why this model was superseded by language. The musical model goes back to Charles Darwin. Darwin assumed that bird songs and language shared a common evolutionary root. Darwin already recognized the multiple components of language. The model is again gaining recognition, but cannot explain the emergence of semantics within melodies. Music with instruments can be traced back about 40,000 years in Homo sapiens. All three of the above models may have analogous or convergent origins; in the first case a protoform arose once, in the second case several times independently.
Questions and answers
Q: What is the study of language called?
A: The study of language is called linguistics.
Q: How do humans communicate with language?
A: Humans communicate with language by using syntax, a set of rules for connecting words together to make statements and questions.
Q: What other forms of communication are used by animals besides language?
A: Other animals may communicate through other means such as inheriting a set of calls which have pre-set functions.
Q: How can human language be changed?
A: Human language can be changed by adding new words, for example, to describe new things.
Q: Are all forms of human communication considered to be language?
A: No, not all forms of human communication are considered to be language; some forms are considered non-verbal communication.
Q: Does UNESCO report that any languages are at risk of becoming extinct?
A: Yes, UNESCO reports that 2,500 languages are at risk of becoming extinct.
Q: Is sign language only used by deaf people who cannot hear?
A: Yes, sign language is primarily used by deaf people who cannot hear.
Related articles
Author
AlegsaOnline.com Language: human communication, structure, history and uses Leandro Alegsa
URL: https://en.alegsaonline.com/art/55911
Sources
- bbc.co.uk : "BBC Languages"