Alternative splicing: how one gene yields multiple mRNA and protein isoforms
Overview of alternative splicing: mechanism, common forms, regulation, biological importance, disease links and quality control in eukaryotes.
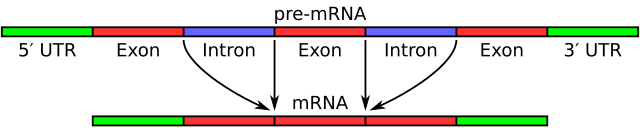
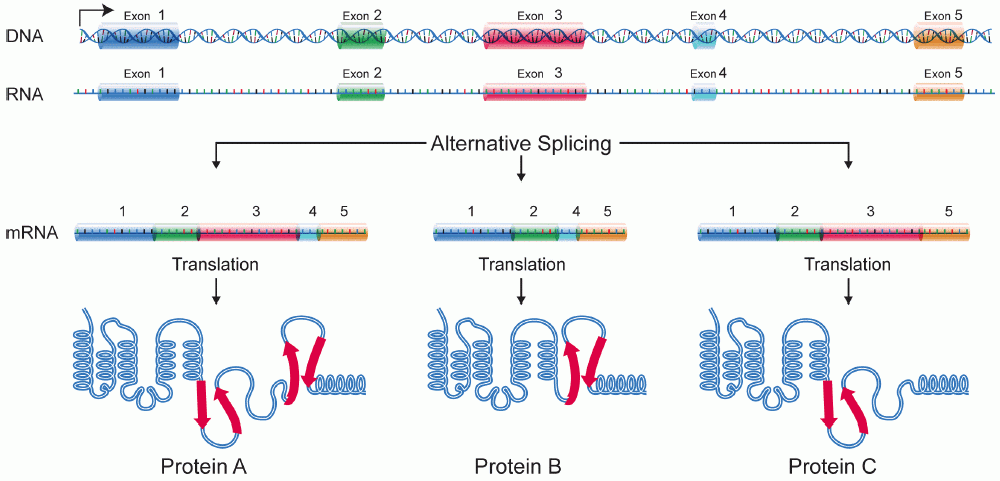
Alternative splicing is a regulated process in which a single gene can give rise to multiple messenger RNA (mRNA) variants and therefore multiple protein products. During gene expression the initial transcript contains both exons (coding segments) and introns (noncoding segments); by selecting different combinations of exons, cells produce distinct mature mRNAs that are later translated into different proteins. This capacity expands the functional repertoire of genomes without increasing gene number.
Image gallery
10 Images





Mechanism
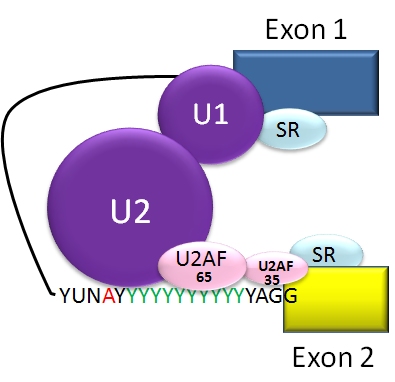
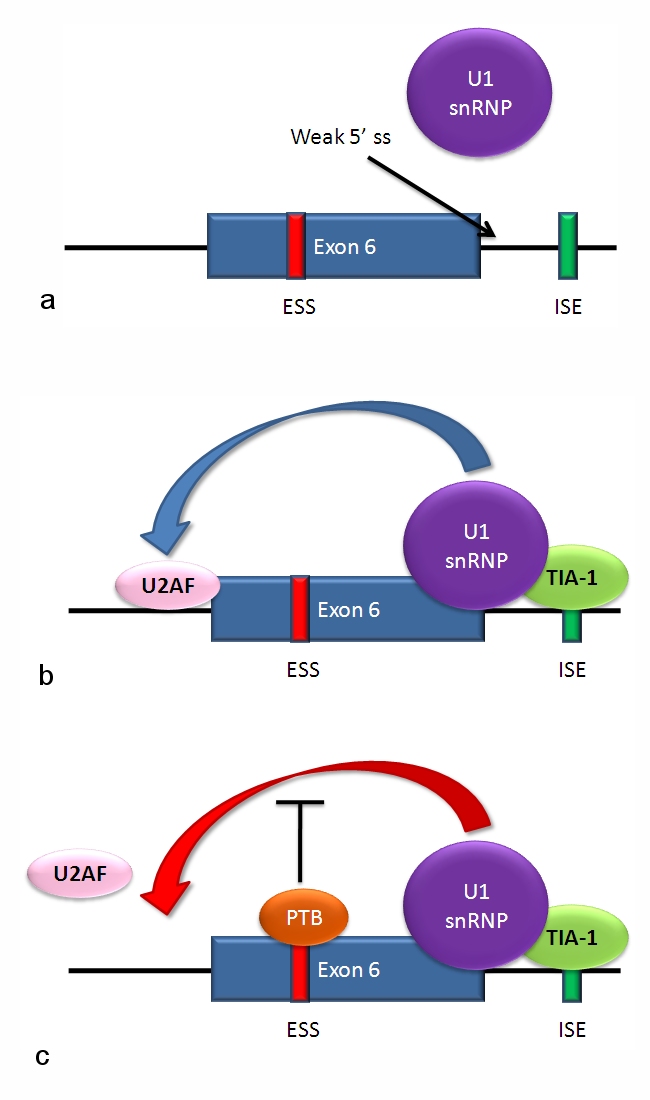
Alternative splicing occurs during the processing of the primary RNA transcript after transcription and as part of the broader process of RNA splicing. The spliceosome, a large ribonucleoprotein complex, recognizes short sequence cues at exon–intron boundaries and catalyzes removal of introns and ligation of exons. Decisions about which splice sites are used depend on sequence motifs in the RNA (cis-elements) and on proteins or RNAs that bind them (trans-acting factors).
Common patterns
There are several recurring ways splice patterns are varied; these produce predictable types of mRNA isoforms. Typical categories include:
- Exon skipping (cassette exons) — an exon may be included or omitted.
- Mutually exclusive exons — two alternative exons are never included together.
- Alternative 5' or 3' splice sites — change the exon boundary and thus coding frame or UTR length.
- Intron retention — an intron remains in the mature RNA and may affect translation or stability.
- Alternative promoters or polyadenylation — produce different first or last exons and modify regulatory regions.
Regulation
Splicing is controlled by activator and repressor proteins that bind enhancers and silencers within exons or introns. Families of factors such as SR proteins and heterogeneous nuclear ribonucleoproteins (hnRNPs) often promote or inhibit particular splice sites. Splicing choices are also influenced by the speed of transcription, chromatin structure and RNA secondary structure, allowing integration of developmental, tissue-specific and environmental signals.
Biological importance and examples
Alternative splicing is widespread in eukaryotes, notably in animals: most human multi-exon genes produce multiple splice isoforms, contributing to cell-type specialization, developmental transitions and adaptability of signaling pathways. Different isoforms can alter enzymatic activity, subcellular localization, interaction partners or regulation by other molecules, so splicing provides a versatile means of modulating protein function.
Clinical relevance and quality control
Errors in splicing or mutations that affect splice sites underlie many human genetic disorders and can contribute to cancer by creating aberrant protein variants. Cells use surveillance pathways such as nonsense-mediated decay to eliminate transcripts with premature stop codons produced by faulty splicing. Therapeutic approaches that modify splicing — for example antisense oligonucleotides that mask or restore splice sites — are an active area of medical research.
Origins and ongoing research
The recognition that single genes can yield multiple proteins emerged with the discovery of split genes and introns in the late 20th century; since then high-throughput RNA sequencing has revealed far greater complexity and context-dependence than previously appreciated. Current research maps tissue-specific splice programs, deciphers the regulatory code, and seeks to exploit splicing modulation for diagnostics and treatments.
Questions and answers
Q: What is alternative splicing?
A: Alternative splicing is a process in which the exons of pre-messenger RNA produced by transcription are reconnected in different ways during RNA splicing, resulting in different mature messenger RNAs from the same gene that get translated into different proteins.
Q: How common is alternative splicing?
A: Alternative splicing is normal in eukaryotes and greatly increases the diversity of proteins that can be encoded by the genome. In humans, around 95% of multiexonic genes are alternatively spliced.
Q: What are some types of alternative splicing?
A: The most common type of alternative splicing is exon skipping, where an exon may be included or omitted from mRNA depending on certain conditions or tissues. There are also other types such as splicing activators and repressors that promote or reduce the use of particular sites respectively, as well as new types being discovered.
Q: How does abnormal variation in alternative splicing affect health?
A: Abnormal variations in alternative splicing can lead to genetic disorders and contribute to cancer development. Non-working products resulting from this process are usually dealt with by post-transcriptional quality control, where they get chopped up by enzymes.
Q: What kind of proteins can be created through alternative splicing?
A: Through alternative splicing, a single gene may code for multiple proteins which increases the diversity of proteins that can be encoded by the genome.
Q: What happens if non-working products result from alternate Spliceing?
A: If non-working products result from alternate Spliceing then they will usually be dealt with by post-transcriptional quality control, where they get chopped up by enzymes.
Related articles
Author
AlegsaOnline.com Alternative splicing: how one gene yields multiple mRNA and protein isoforms Leandro Alegsa
URL: https://en.alegsaonline.com/art/3063
Sources
- doi.org : 10.1146/annurev.biochem.72.121801.161720
- pubmed.ncbi.nlm.nih.gov : 12626338
- doi.org : 10.1038/ng.259
- pubmed.ncbi.nlm.nih.gov : 18978789
- doi.org : 10.1038/nrm1645
- pubmed.ncbi.nlm.nih.gov : 15956978
- doi.org : 10.1101/gad.1643108
- pubmed.ncbi.nlm.nih.gov : 18245441
- doi.org : 10.1016/j.biocel.2007.02.016
- pubmed.ncbi.nlm.nih.gov : 17416541
- ncbi.nlm.nih.gov : "A global view of cancer-specific transcript variants by subtractive transcriptome-wide analysis"
- doi.org : 10.1371/journal.pone.0004732
- pubmed.ncbi.nlm.nih.gov : 19266097
- ncbi.nlm.nih.gov : "Aberrant RNA splicing and its functional consequences in cancer cells"