Cryptomonads (Cryptophyta): small flagellate algae with complex plastids
Cryptomonads are mostly unicellular, flagellated algae found in fresh, brackish, and marine waters. Many contain complex chloroplasts with a nucleomorph and can be photosynthetic, mixotrophic, or heterotrophic.
Overview
Cryptomonads, also called Cryptophyta, are a group of mostly unicellular, flagellated protists commonly treated as a phylum of algae. Individuals are typically flattened and measure roughly 10–50 µm across. They occur worldwide in freshwater systems and in marine and brackish environments, where they often contribute to planktonic communities and primary production.
Image gallery
3 Images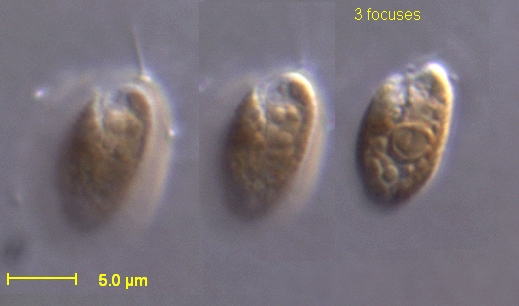


Morphology and distinctive features
Cryptomonads characteristically bear an anterior groove or pocket from which two slightly unequal flagella emerge. The flagella often have fine hairs and help with swimming and feeding. Cells commonly possess ejectisomes (extrusome-like organelles) that can release threads in response to disturbance. Most species contain chloroplasts with accessory pigments such as phycobiliproteins that give some species distinctive colors (blue-green, brown, or red hues).
Plastids, nucleomorph and origins
The plastids of many cryptomonads are the product of secondary endosymbiosis: a eukaryotic host incorporated a red algal cell. Remnants of that red algal nucleus — a tiny, reduced nucleus called a nucleomorph — are retained between the plastid membranes in some lineages. This arrangement (plastids surrounded by four membranes plus a nucleomorph) makes cryptomonads important models for studying endosymbiotic events and genome reduction.
Ecology, nutrition and lifecycle
Cryptomonads are abundant in lakes, rivers and coastal waters, and play an important role as primary producers and as food for zooplankton and protozoan grazers. While many species are obligate photoautotrophs, several are mixotrophic: they combine photosynthesis with ingestion of particulate food or dissolved organic material. Reproduction is usually by simple cell division; clearly documented sexual cycles are rare or poorly known in this group.
Importance, research and examples
Genera commonly encountered in ecological and laboratory studies include Cryptomonas, Guillardia and Hemiselmis. The genome and nucleomorph from species such as Guillardia have been sequenced and provided key insights into secondary endosymbiosis and gene transfer. Cryptomonads are studied in ecology, evolution and biotechnology—for example, their pigments have applications as fluorescent markers and in research on photosynthesis.
Notable distinctions
- Four-membrane plastids with a nucleomorph (in many species).
- Anterior pocket and two unequal flagella for locomotion.
- Presence of ejectisomes for rapid response to threats.
- Varied nutrition: autotrophy, mixotrophy and occasional phagotrophy.
- Found across freshwater, brackish and marine habitats.
For overviews of classification and diversity consult taxonomic summaries and collections of ecological studies: see a general taxonomy entry at classification resources, reviews of aquatic distribution at habitat surveys, technical material on plastid structure, and detailed descriptions of flagellar anatomy at flagellar studies.

Systematics of the Cryptophyceae
The systematic classification of the Cryptophyceae into different genera was mainly based on morphological characteristics and pigmentation.
- One of the most important features here is the periplast. The periplast is a layered cell envelope -- cryptophyceae do not form a cell wall -- consisting of an inner and an outer periplast component made of proteins. Between them is the cell's plasma membrane. Both periplast components show very fine structuring. For example, the inner periplast component may consist of polygonal plates, overlapping rectangular plates, or a continuous layer. The outer periplast component may also be composed of plates or of rosette scales and fine fibrils.
- All Cryptophyceae have a cell invagination, which is lined with explosive organelles, the so-called ejecto- or ejectisomes (see extrusome). The opening of this cell invagination can either be small with a blind ending sac behind it (pharynx) or elongated following the cell invagination (furrow). Combinations of furrow and pharynx are also possible.
- In some genera, the nucleomorph is not freely located in the periplastid space, but is embedded in the pyrenoid matrix (spatially separated by the two inner envelope membranes of the plastid).
- Of the phycobilisomes, the light-collecting complexes of red algae (and glaucocystophyceae and cyanobacteria), which originally contained three different blue or red pigments, only phycoerythrin remained in the Cryptophyceae. In the Cryptophyceae, however, seven different types of phycoerythrin evolved from the original red phycoerythrin, four of which are blue in color and are therefore called phycocyanins, although they are not directly related to the true phycocyanins from the phycobilisomes.
- The microtubular flagellar roots, with which the flagella are anchored in the cells, also show differences.
From the combination of the different characteristics
- Structure of the periplast,
- Shape of the cell invagination,
- Position of the nucleomorph,
- Pigment type and
- Structure of the flagellar root apparatus
result in the different genera.
However, research into the relationships within the Cryptophyceae using methods of molecular phylogenetic analysis (= creation of phylogenetic trees based on DNA sequences), revealed a much more complex picture. Cryptophyceae are probably dimorphic, i.e. they can form two different cell types. Therefore, two cell forms of one genus were probably mistaken for two different genera several times. Dimorphism has been reliably demonstrated in the genera Proteomonas and Cryptomonas . Even a leucoplast, a colorless plastid that has lost the ability to photosynthesize, is not a sure characteristic of a distinct genus. The former genus Chilomonas proved to be a colorless Cryptomonas, of which, moreover, at least three different evolutionary lineages exist within Cryptomonas. The remaining genera of the Cryptophyceae probably also require a revision of their systematics.
Genera of Cryptophyceae according to preliminary research: Chroomonas, Cryptomonas (contains the formerly independent genera Campylomonas and Chilomonas), Geminigera, Goniomonas, Guillardia, Hanusia, Hemiselmis, Komma, Plagioselmis, Proteomonas, Rhinomonas, Rhodomonas (Pyrenomonas), Teleaulax.

Questions and answers
Q: What are Cryptomonads?
A: Cryptomonads are a phylum of algae that are commonly found in freshwater and sometimes in marine and brackish habitats.
Q: Do Cryptomonads have chloroplasts?
A: Yes, most Cryptomonads have chloroplasts.
Q: How big is each Cryptomonad cell?
A: Each Cryptomonad cell is around 10-50 μm in size and flattened in shape.
Q: What kind of groove or pocket does a Cryptomonad cell have?
A: A Cryptomonad cell has an anterior groove or pocket.
Q: How many flagella do Cryptomonads typically have?
A: Cryptomonads typically have two slightly unequal flagella at the edge of the anterior pocket.
Q: Do Cryptomonads exhibit mixotrophy?
A: Yes, some Cryptomonads exhibit mixotrophy, which means they use mixed sources of energy.
Q: Where are Cryptomonads commonly found?
A: Cryptomonads are commonly found in freshwater environments, and sometimes in marine and brackish habitats.
Related articles
Author
AlegsaOnline.com Cryptomonads (Cryptophyta): small flagellate algae with complex plastids Leandro Alegsa
URL: https://en.alegsaonline.com/art/24460
Sources
- nar.oxfordjournals.org : "Lateral transfer of introns in the cryptophyte plastid genome"
- doi.org : 10.1093/nar/gkn095
- ncbi.nlm.nih.gov : 2396441
- pubmed.ncbi.nlm.nih.gov : 18397952
- life.umd.edu : "Cryptophyta - the cryptomonads"